Gerridae
 | ||
|---|---|---|
 Aquarius remigis adulto | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Arthropoda | |
| Clase: | Insecta | |
| Orden: | Hemiptera | |
| Suborden: | Heteroptera | |
| Infraorden: | Gerromorpha | |
| Superfamilia: | Gerroidea | |
| Familia: | Gerridae Leach, 1815 | |
| Subfamilias[1] | ||
| ||
[editar datos en Wikidata] | ||
Los Guérridos (Gerridae) son una familia de insectos del orden Hemiptera y del suborden Heteroptera, conocidos popularmente como Zapateros, Chinches patinadoras, Zancudos de agua, Patinadores de agua o simplemente Patinadores. Sus piezas bucales son de tipo perforador-succionador, y es característica su habilidad para deslizarse sobre el agua, distribuyendo su peso para aprovechar la tensión superficial de la superficie del agua, por lo que son fácilmente observables en ríos y lagos. Se han identificado más de 1700 especies de guérridos, de los cuales un 10% son especies marinas.[2]
A pesar de que el 90% de los guérridos viven en aguas continentales, el género marino Halobates convierte a la familia Gerridae en un caso bastante extraordinario. El género Halobates fue estudiado con detenimiento por primera vez entre 1822 y 1883 cuando Francis Buchanan White recogió varios ejemplares de distintas especies durante la Expedición Challenger.[3] Por aquel entonces, Eschscholtz descubrió tres especies de Gerridae, lo que atrajo atención sobre esta familia a pesar del escaso conocimiento sobre su biología.[3] Desde entonces, los guérridos se han seguido estudiando por su capacidad de patinar sobre el agua y por sus interacciones sociales. Algunos Gerridae de pequeño tamaño se han confundido con otro tipo de insectos, los Veliidae; el rasgo más definitorio para distinguirlos se halla en las diferencias internas entre sus aparatos reproductores, por lo que es prácticamente imposible hacerlo a simple vista y se precisan tanto una formación específica como instrumentos de análisis adecuados.
La familia Gerridae se caracteriza por sus pelos hidrófobos, sus garras retráctiles preapicales y su cuerpo y patas esbeltos.[4] Los pelos hidrófobos son pelos minúsculos que contienen más de mil micropelos por milímetro.[4] Todo su cuerpo está cubierto por estos pelos, lo que les da resistencia frente a salpicaduras y gotas de agua.
Descripción
La familia Gerridae se caracteriza físicamente por tener pelos hidrófugos, garras preapicales retráctiles y patas y cuerpo alargados.[4]
Los pelos hidrófugos son pequeños micropelos hidrófobos. Se trata de pelos diminutos con más de mil micropelos por mm.[4] Todo el cuerpo está cubierto por estos pelos, lo que proporciona al zancudo acuático resistencia a las salpicaduras o gotas de agua. Estos pelos repelen el agua, evitando que las gotas pesen sobre el cuerpo.
Tamaño
Generalmente son insectos pequeños, de patas largas y la longitud del cuerpo de la mayoría de las especies está entre 2 y 12 mm. Algunas tienen entre 12 y 25 mm.[5] Entre los géneros más extendidos, el del hemisferio norte Aquarius incluye las especies más grandes, que generalmente superan los 12 mm (0,5 plg), al menos entre las hembras, y las especies más grandes con una media de unos 24 mm (0,9 plg). [5][6] Las hembras suelen ser más grandes que los machos de su propia especie,[5] pero parece ser al revés en la especie más grande, la relativamente poco conocida Gigantometra gigas de los arroyos del norte de Vietnam y el sur de China adyacente. Suele alcanzar una longitud corporal de unos 36 mm (1,42 plg) en los machos sin alas y de 32 mm (1,26 plg) en las hembras aladas (los machos alados, sin embargo, sólo son en promedio ligeramente más grandes que las hembras). En esta especie, cada pata media y trasera puede superar los 10 cm (4 plg).[7]
Antenas

Los zancudos tienen dos antenas con cuatro segmentos cada una. Los segmentos antenales están numerados del más cercano a la cabeza al más lejano. Las antenas tienen cerdas cortas y rígidas en el segmento III.[8] Las longitudes relativas de los segmentos de las antenas pueden ayudar a identificar especies únicas dentro de la familia Gerridae, pero en general, el segmento I es más largo y corpulento que los tres restantes.[9] Los cuatro segmentos combinados no suelen ser más largos que la longitud de la cabeza del estrícido acuático.
Tórax
El tórax de los zancudos es generalmente largo, estrecho y de pequeño tamaño. Suele medir entre 1,6 y 3,6 mm de longitud según la especie, con algunos cuerpos más cilíndricos o redondeados que otros.[9] El pronoto, o capa externa del tórax, de los zancudos puede ser brillante u opaco según la especie, y estar cubierto de micropelos para ayudar a repeler el agua. [8] El abdomen de un estrícido acuático puede tener varios segmentos y contiene tanto el metasternum como el omphalium.[8]
Apéndices
Los Gerridae tienen patas delanteras, medias y traseras. Las patas delanteras son más cortas y tienen garras preapicales adaptadas para pinchar a las presas. Las garras preapicales son garras que no están al final de la pata, sino a medio camino, como las mantis. Las patas medias son más largas que el primer par y más cortas que el último y están adaptadas para la propulsión a través del agua. El par posterior es el más largo y sirve para distribuir el peso sobre una gran superficie, así como para dirigir al insecto por la superficie del agua. Las patas delanteras están unidas justo después de los ojos, mientras que las centrales están más cerca de las traseras, que se unen a la mitad del tórax pero se extienden más allá del extremo terminal del cuerpo.[8]
Géneros de Gerridae
- Aquarius (Schellenberg, 1800).
- Gerris (Fabricius, 1794).
- Halobates (Eschscholtz, 1822).
- Limnogonus (Stal, 1868).
- Limnoporus (Stal, 1868).
- Metrobates (Uhler, 1878).
- Neogerris (Matsumara, 1913).
- Rheumatobates (Bergroth, 1892).
- Trepobates (Uhler, 1883).
Especies de Europa
- Aquarius cinereus (Puton, 1869)
- Aquarius najas (De Geer, 1773)
- Aquarius paludum (Fabricius, 1794)
- Aquarius ventralis (Fieber, 1860)
- Gerris argentatus (Schummel, 1832)
- Gerris brasili (Poisson, 1941)
- Gerris costae (Herrich-Schaeffer, 1850)
- Gerris gibbifer (Schummel, 1832)
- Gerris lacustris (Linnaeus, 1758)
- Gerris maculatus (Tamanini, 1946)
- Gerris odontogaster (Zetterstedt, 1828)
- Gerris sphagnetorum (Gaunitz, 1947)
- Gerris thoracicus (Schummel, 1832)
- Gerris asper (Fieber, 1860)
- Gerris lateralis (Schummel, 1832)
- Limnoporus rufoscutellatus (Latreille, 1807)
Evolución
El Cretogerris, procedente del ámbar Charentese del Cretácico (Albiano) de Francia, se sugirió inicialmente como un gérrido.[10], aunque posteriormente se interpretó como un miembro indeterminado de Gerroidea. Son morfológicamente similares a los Chresmoda, un enigmático género de insectos conocido desde el Jurásico Tardío hasta el Cretácico Medio con un estilo de vida presumiblemente similar.
Polimorfismo en las alas
El polimorfismo alar (es decir, la presencia de múltiples morfos alares en una especie dada) ha evolucionado de forma independiente en múltiples ocasiones en Gerridae, así como la pérdida completa de las alas,[11] algo que ha sido importante para la evolución de la variedad en las especies que vemos hoy en día, y la dispersión de Gerridae. La existencia de polimorfismo alar en una especie determinada puede explicarse como un caso particular de síndrome de oogénesis-vuelo. Siguiendo este razonamiento, que se aplica comúnmente en los insectos, el desarrollo de alas cortas proporciona al individuo la capacidad de dedicar las reservas de energía que normalmente se utilizarían para el desarrollo de las alas y de los músculos alares a aumentar la producción de huevos y a reproducirse precozmente, lo que en última instancia mejora la capacidad del individuo.[12] La capacidad de que una cría tenga crías con alas y la siguiente no, permite a los zancudos acuáticos adaptarse a entornos cambiantes. Dependiendo del entorno y de la estación del año, se necesitan alas largas, medianas, cortas o inexistentes. Las alas largas permiten volar a una masa de agua vecina cuando hay demasiada gente, pero pueden mojarse y pesar. Las alas cortas pueden permitir desplazamientos cortos, pero limitan lo lejos que puede dispersarse un gérrido. El no tener alas evitan que un gérrido sea pesado, pero impiden su dispersión.
El polimorfismo de las alas es común en los gérridos, a pesar de que la mayoría de las poblaciones univoltinas (una nidada) son completamente ápteras (sin alas) o macrópteras (con alas).[13] Las poblaciones ápteras de gérridos estarían restringidas a hábitats acuáticos estables que experimentan pocos cambios en el medio ambiente, mientras que las poblaciones macrópteras pueden habitar aguas más cambiantes y variables.[13] Las aguas estables suelen ser grandes lagos y ríos, mientras que las aguas inestables suelen ser pequeñas y estacionales. Los gérridos producen formas aladas con fines de dispersión y los individuos macrópteros se mantienen debido a su capacidad para sobrevivir en condiciones cambiantes.[13] Las alas son necesarias si es probable que la masa de agua se seque, ya que el gérrido debe volar hasta una nueva fuente de agua. Sin embargo, las formas sin alas se ven favorecidas debido a la competencia por el desarrollo de los ovarios y las alas, y el éxito reproductivo es el objetivo principal debido a la teoría del gen egoísta. Los gérridos invernantes suelen ser macrópteros, o con alas, para poder volar de vuelta a su hábitat acuático después del invierno. Un mecanismo de cambio ambiental controla el dimorfismo estacional observado en las especies bivoltinas, o especies que tienen dos nidadas al año.[13] Este mecanismo de cambio es el que ayuda a determinar si evolucionará o no una nidada con alas. La temperatura también desempeña un papel importante en el cambio fotoperiódico.[13] La temperatura marca las estaciones y, por tanto, cuándo se necesitan las alas, ya que hibernan durante el invierno. En última instancia, estos mecanismos de cambio alteran los alelos genéticos para las características de las alas, ayudando a mantener la dispersión biológica.
Naturaleza de la capacidad de caminar sobre el agua

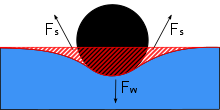
Los zancudos acuáticos son capaces de caminar sobre el agua debido a una combinación de varios factores. Utilizan la alta tensión superficial del agua y sus largas patas hidrófobas para mantenerse sobre el agua. Las especies de Gerridae aprovechan esta tensión superficial gracias a sus patas altamente adaptadas y a la distribución de su peso.
Las patas son largas y delgadas, lo que permite distribuir el peso del cuerpo sobre una gran superficie. Las patas son fuertes, pero tienen una flexibilidad que les permite mantener su peso uniformemente distribuido y fluir con el movimiento del agua. Los pelos hidrófugos recubren la superficie del cuerpo del zancudo acuático. Hay varios miles de pelos por milímetro cuadrado, lo que proporciona al zancudo acuático un cuerpo hidrófugo que evita la humedad de las olas, la lluvia o el rocío, lo que podría inhibir su capacidad para mantener todo su cuerpo por encima de la superficie del agua si el agua se atascara y pesara sobre el cuerpo.[4] Esta postura de mantener la mayor parte del cuerpo por encima de la superficie del agua se denomina posición epipleustónica, que es una característica definitoria de los zancudos acuáticos. Si el cuerpo del zancudo se sumergiera accidentalmente, por ejemplo por una gran ola, los diminutos pelos atraparían el aire. Diminutas burbujas de aire en todo el cuerpo actúan como flotabilidad para llevar al zancudo de agua a la superficie de nuevo, mientras que también proporciona burbujas de aire para respirar bajo el agua.[4] A pesar de su éxito en la superación de la inmersión en el agua, sin embargo, los zancudos de agua no son tan competentes en petróleo, y derrames de petróleo experimentales de petróleo han sugerido que el petróleo derramado en sistemas de agua dulce puede provocar la inmovilidad y la muerte de los zancudos acuáticos.[14]
Los diminutos pelos de las patas proporcionan tanto una superficie hidrófoba como una mayor superficie para repartir su peso sobre el agua. Las patas medias utilizadas para remar tienen pelos en la tibia y el tarso particularmente bien desarrollados para ayudar a aumentar el movimiento a través de la capacidad de empuje.[4] El par de patas traseras se utilizan para la dirección [15]Cuando comienza la brazada de remo, los tarsos medios de los gérridos se presionan rápidamente hacia abajo y hacia atrás para crear una onda superficial circular en la que la cresta puede utilizarse para impulsar un empuje hacia delante. [4] La onda semicircular creada es esencial para la capacidad del zancudo acuático de moverse rápidamente, ya que actúa como una fuerza contra la que empujar. Como resultado, los zancudos acuáticos a menudo se mueven a 1 metro por segundo o más rápido. [16]
Referencias
- ↑ Schuh R.T., Slater J.A. (1995). True Bugs of the World (Hemiptera: Heteroptera). Classification and Natural History. Cornell University Press, Ithaca, New York, USA. 336 pp.
- ↑ Lancaster, J.B.; Briers, R., eds. (2008). Aquatic insects: challenges to populations. CABI. pp. 23, 270, 284.
- ↑ a b Cheng, L. (1985). Biology of Halobates «Biology of Halobates (Heteroptera: Gerridae)». Annual Review of Entomology 30 (1): 111–135. doi:10.1146/annurev.en.30.010185.000551.
- ↑ a b c d e f g h Ward, J.V. (1992). Aquatic Insect Ecology: 1. Biology and habitat. New York: Wiley & Sons. pp. 74, 96, 172, 180.
- ↑ a b c Andersen, N.M. (1997). «Un análisis filogenético de la evolución del dimorfismo sexual y los sistemas de apareamiento en los zancudos acuáticos (Hemiptera: Gerridae)». Biological Journal of the Linnean Society 61 (3): 345-368. doi:10.1006/bijl.1996.0130.
- ↑ Damsgaard, J.; Zettel, H. (2003). «Diversidad genética, filogenia de especies y biogeografía histórica del grupo Aquarius paludum (Heteroptera: Gerridae)». Insect Systematics & Evolution 34 (3): 313-328. doi:10.1163/187631203788964791.
- ↑ Tseng, M.; Rowe, L. (1999). «Dimorfismo sexual y alometría en el trancón Gigantometra gigas». Canadian Journal of Zoology 34 (6): 923-929. S2CID 56016772. doi:10.1139/z99-071.
- ↑ a b c d Merrit, R.; Cummins, K. (1996). Una introducción a los insectos acuáticos de Norteamérica. Kendall/Hunt Pub. Co. pp. 275-282.
- ↑ a b Slater, J (1995). Bichos Verdaderos del Mundo (Hemiptera: Heteroptera). Comstock Pub. Associates. pp. 1-15.
- ↑ Perrichot, Vincent; Nel, André; Neraudeau, Didier (October 2005). «Gerromorphan bugs in Early Cretaceous French amber (Insecta: Heteroptera): first representatives of Gerridae and their phylogenetic and palaeoecological implications». Cretaceous Research (en inglés) 26 (5): 793-800. doi:10.1016/j.cretres.2005.05.003.
- ↑ Armisén, David; Viala, Séverine; Cordeiro, Isabelle da Rocha Silva; Crumière, Antonin Jean Johan; Hendaoui, Elisa; Bouquin, Augustin Le; Duchemin, Wandrille; Santos, Emilia; Toubiana, William; Vargas-Lowman, Aidamalia; Floriano, Carla Fernanda Burguez; Polhemus, Dan A.; Wang, Yan-hui; Rowe, Locke; Moreira, Felipe Ferraz Figueiredo (10 de enero de 2022). Transcriptome-based phylogeny of the semi-aquatic bugs (Hemiptera: Heteroptera: Gerromorpha) reveals patterns of lineage expansion in a series of new adaptive zones (en inglés). pp. 2022.01.08.475494. doi:10.1101/2022.01.08.475494.
- ↑ Roff, Derek A. (December 1990). «The Evolution of Flightlessness in Insects». Ecological Monographs (en inglés) 60 (4): 389-421. ISSN 0012-9615. doi:10.2307/1943013.
- ↑ a b c d e Koga, Hayashi. 1991. Territorial behavior of both sexes in the water strider Metrocoris histrio (Hemiptera: Gerridae) during the mating season. Journal of Insect Behavior, Volume 6 (1).
- ↑ Black, Tyler (December 2019). «Los efectos de un derrame simulado de betún diluido en invertebrados en un entorno de lago boreal». Tesis del MSC.
- ↑ Williams, D.; Feltmate, B. (1992). Insectos acuáticos. CAB International. pp. 48, 121, 218. ISBN 0-85198-782-6.
- ↑ Andersen, Nils Moller; Cheng, Lanna (2004). «The marine insect Halobates (Heteroptera: Gerridae): Biology, Adaptations, Distribution and Phylogeny». Oceanography and Marine Biology: An Annual Review 42: 119-180. doi:10.1201/9780203507810.ch5. Archivado desde el original el 20 de agosto de 2011.
Enlaces externos
- «Gerridae (TSN 103801)». Sistema Integrado de Información Taxonómica (en inglés).
- Imágenes en gran formato. Variedades aladas y ápteras
- Vídeo del movimiento sobre el agua
| Control de autoridades |
|
|---|
 Datos: Q270322
Datos: Q270322 Multimedia: Gerridae / Q270322
Multimedia: Gerridae / Q270322 Especies: Gerridae
Especies: Gerridae







